|
Bartonellae are highly adaptive organisms that have the ability to evade the host immune system and cause persistent bacteraemia by occupying the
host’s erythrocytes. Bartonella spp. is under-studied and health care professionals often misdiagnose Bartonella-related
infections. The aim of this study was to investigate the carriage of Bartonella spp. circulating in human and animal populations in
Gauteng using culturing and polymerase chain reaction (PCR) detection. A total of 424 human, 98 cat, 179 dog, and 124 wild rodent blood
samples were plated onto specialised media and incubated for 7–21 days at 37 ºC in CO2. Culture isolates
morphologically similar to Bartonella control strains were confirmed by PCR and sequenced to determine species. Deoxyribonucleic
acid (DNA) was extracted from all blood samples and tested by nested PCR. Bartonella could only be cultured from the cat and rodent
specimens. Cat isolates were > 99% similar to Bartonella henselae URBHLIE 9, previously isolated from an endocarditis patient, and
rat isolates were > 98% similar to either RN24BJ (candidus ‘Bartonella thailandensis’) or RN28BJ, previously isolated from
rodents in China. The PCR prevalences were 22.5% in HIV-positive patients, 9.5% in clinically healthy volunteers, 23.5% in cats, 9% in dogs and 25%
in rodents. Findings of this study have important implications for HIV-positive patients.
Background
Bartonella fall within the alpha-2 subgroup of the class Proteobacteria (Jacomo, Kelly & Raoult 2002). Recent studies have indicated
that Bartonella species (spp.) have some degree of relatedness to other alpha-2 Proteobacteria including Brucella species,
Afipia species, Agrobacterium tumefaciens, Bradyrhizobium species, and Bosea species (Duncan, Maggi &
Breitschwerdt 2007; Greub & Raoult 2002; Houpikian & Raoult 2001; Jacomo et al. 2002; Pretorius, Beati & Birtles 2004;
Rolain et al. 2004). Current knowledge suggests that there are more than 20 species and subspecies included within this genus
(Márquez et al. 2008). Approximately 13 species have been associated with human diseases (Pérez-Martínez et al.
2009; Maggi et al. 2009; Pons et al. 2008) affecting both immunocompetent and immunocompromised individuals. At least six
species affecting humans have been isolated from domestic cats and dogs (Chomel et al. 2006).Bartonellae are pleomorphic, fastidious, oxidase-negative, Gram-negative bacilli (Jacomo et al. 2002; Maurin & Raoult 1996).
Bartonella species growth is hemin-dependent (Wong et al. 1995), therefore the addition of hemin-rich rabbit blood or horse blood to
the agar yields better growth than sheep blood (Jacomo et al. 2002). Growth occurs on enriched medium at 37 °C with 5% carbon dioxide
(CO2); however, growth also occurs in broth with fetal bovine serum, and in various tissue culture systems including cell lines (La
Scola & Raoult 1999). On average primary isolates appear after 12 to 14 days (Jacomo et al. 2002), although it has been reported that
primary isolation can take up to 45 days (Maurin et al. 1994). Subcultured colonies have been found to appear after only three to five
days (Jacomo et al. 2002).
Objectives
In this study, human immunodeficiency virus (HIV) positive people, clinically healthy volunteers, impounded cats, impounded dogs, and commensal
rodents were investigated for carriage of Bartonella spp. The objective was to determine the rates of infection in the study groups and to
determine the species responsible for infection in the study populations.
Literature review
Various studies have been carried out globally to determine the prevalence of Bartonella spp. in humans and animals. There is very little
data available for the culture or molecular prevalence of human Bartonella. The lowest prevalence reported in humans was 3% and the highest
was 33.3%, in the USA (Breitschwerdt et al. 2007; Bonilla et al. 2009; Koehler et al. 2003). The prevalence in cat populations
was from as little as 2% in Canada (Kamrani et al. 2008) to as high as 53% in France (Heller et al. 1997), in dog populations from 0%
in the United Kingdom (Birtles et al. 2002) to 16% in Korea (Kim et al. 2009), and in rodent populations from 6% in Indonesia
(Winoto et al. 2005) to 43.5% in China (Ying et al. 2002). The prevalence of Bartonella spp. is largely unknown in South Africa. There are some case reports of bacillary angiomatosis caused by
Bartonella spp. in HIV positive patients (Frean, Arndt & Spencer 2002). A non-random pilot survey of outpatients of three Johannesburg
HIV clinics has been done on a relatively small sample group (n = 188 patients). This study showed a 10% prevalence
of Bartonella henselae in blood of HIV-positive patients, determined by nested polymerase chain reaction
(PCR) (Frean et al. 2002). A prevalence study in domestic and wild felines in southern Africa published by Kelly et al. (1996)
reported 23% (n = 171) prevalence. A study of rodents in South Africa showed a high rate of infection (44%) with a wide range of subtypes
of bartonellae (Pretorius et al. 2004).
Sample collection
Ethical considerations
Ethical approval for collection of samples from humans and animals was granted by the Human Research Ethics Committee (Medical) of the University
of the Witwatersrand (Ethics Approval Number: M070637), the National Health Laboratory Service Animal Ethics Committee (Ethics Approval Number:
2007/112), and the University of the Witwatersrand Animal Ethics Screening Committee (AESC) (Ethics Approval Number: 2008/56/03).
Recruitment of participants and informed consent
A convenience sample of 382 HIV-positive patients was recruited from the Chris Hani Baragwanth Hospital (Soweto, Gauteng), HIV-clinic. HIV-status
was known as the clinic only treats HIV-positive patients. Specimen collection started in September 2007 and was completed by the end of January
2008. Survey objectives and consent forms were explained to prospective participants by an available HIV-counselor employed at the clinic, a
laboratory aid or by the phlebotomists drawing the blood samples. Patients were assured that the study was voluntary and results would not
directly benefit nor disadvantage them in any way. Informed consent forms were completed by the participating patients before blood collection.
The recruited volunteers were provided with an information sheet containing the contact details of the Special Bacterial Pathogens Reference
Laboratory (SBPRL) at the National Institute for Communicable Diseases (NICD) to which queries were addressed. Clinically healthy volunteers were recruited from the National Institute for Communicable Diseases and a local animal shelter for blood
collection. Due to ethical considerations, the volunteers were not required to disclose their HIV status, and health was assumed on the basis of
no observable signs of illness at the time of blood collection. These volunteers were also required to complete an informed consent form and
participation was completely voluntary. Two 4 mL tubes of blood were collected from each volunteer, 1 tube containing the anticoagulant
ethylenediaminetetra-acetic acid (EDTA) and the other a plain red-top tube in which the blood was allowed to clot. Blood specimens were stored at
minus 20 °C until use.
Animal samples
Blood was collected from dogs and cats that were undergoing euthanasia, surgery or sterilisation procedures at a local animal shelter. Two 2 mL
tubes of blood (as described above) were collected from each cat and dog. Samples were stored at 4 °C until use.Rodents were prospectively sourced from a pest control company operating in the Ekurhuleni Metropolitan area. Rodents were trapped alive,
anaesthetised using CO2, and exsanguinated by cardiac puncture. Blood was collected using a syringe and needle from each rodent and was
divided into two tubes, one red top and one purple top (as above). Specimens were stored at 4 °C until use.
Procedures
Culturing of Bartonella spp.
All blood samples were plated onto 5% rabbit blood-supplemented Columbia agar (hereafter referred to as Bartonella medium). Culturing was
carried out in a class two biohazard safety cabinet to reduce the risk of contamination. Ethylenediaminetetra-acetic acid blood was predominantly
used for culturing, unless the EDTA specimen was insufficient (especially for cat and dog samples), in which case some of the clotted blood (packed
cells excluding serum) was used. Approximately 100 μL – 150 μL of each whole blood sample was inoculated onto
fresh Bartonella media and spread over the surface using sterile glass spreaders. Inoculated media were incubated in 5% CO2
at 37 °C for seven ± two days before being checked for growth.If small dry colonies, pitting into the media were present, they were Gram stained, and plated out onto fresh media. Media with faint or no
growth were re-incubated as above to a maximum of two months with periodic examinations every seven days. All blood culture isolates were
confirmed by PCR and stored in tryptic soy broth (TSB) with 10% (v/v) glycerol at minus 80 °C.
Genus specific polymerase chain reaction amplification
All culture isolates and blood samples were tested by genus specific PCR amplification of the intergenic spacer (ITS) region between the 16S and
23S rRNA genes for evidence of Bartonella Deoxyribonucleic acid (DNA). Deoxyribonucleic acid extractions were carried out using the QIAamp
DNA mini kit (Qiagen, Germany) as per kit protocol.Published genus-specific primers were selected as shown in Table 1 (Roux & Raoult 1995; Seki et al. 2006). The reverse primer QHVE-14
(Seki et al. 2006) was modified by elongating the 5’ end by three nucleotides to decrease non-specific amplification.
|
TABLE 1:
Published genus-specific primers.
|
Polymerase chain reaction confirmation of cultured isolates was carried out using primers QHVE-1 and QHVE-3, whereas the blood-extracted DNA
required a nested PCR using the QHVE-12 and QHVE-14b inner primers. Table 2 shows the primer binding localities and product sizes of the six most
commonly isolated human Bartonella spp., although other species within the genus are also detectable using these primers.
|
TABLE 2:
Primer binding localities and product sizes of the six most commonly isolated human Bartonella spp.
|
The single or first round PCR amplifications were carried out in a reaction mixture (50 mL) containing 20 ρmol of each primer
(QHVE-1 and QHVE-3), 1 x Buffer II, 2 mM MgCl2, 1.5 U AmpliTaq DNA polymerase (Applied Biosystems, USA), 200 μM of each
deoxyribonucleotide triphosphate (dNTP) (Thermo Scientific, United Kingdom), and 1% (v/v) Triton-X 100 surfactant additive. The amount of DNA was
used depended on the source of the DNA. For the culture DNA, 20 ng of template was included into the reaction, whereas 5 mL of the total blood
extracted extracted DNA was included into the reaction. Polymerase chain reactions were performed on a VERITI Thermocycler (Applied Biosystems)
under the following conditions: 2 min initial denaturation step at 94 °C, followed by 35 cycles of the following steps: denaturation at
94 °C for 30 s, primer annealing at 52 °C for 30 s, and elongation at 72 °C for 60 s. A final elongation step concluded the
amplification at 72 °C for 6 min. Polymerase chain reaction products were maintained at 4 °C until being added to the reaction mixtures of
the nested round. For the nested PCR reactions, a final volume of 50 μL consisted of 2 μL of first-round product, 1 x Buffer II,
1.5 mM MgCl2, 30 ρmol of each inner primer (QHVE-12 and QHVE-14b), 200 μM of each dNTP, and 1.5 U AmpliTaq DNA polymerase.
The reactions were amplified as previously described, with a variation in the
annealing temperature (55 °C).Amplicon analysis was performed on a 2% (w/v) Tris-acetic acid-EDTA (TAE) buffer agarose (WhiteSci, USA) gel, supplemented with 0.5 μg/mL
ethidium bromide. Electrophoresis was carried out at 100 V in 1 x TAE for 40 min. Gels were visualised by ultraviolet illumination.
Deoxyribonucleic acid sequencing
The culture isolates were speciated by sequencing of the ITS region using the outer primers (QHVE-1 and QHVE-3). Amplicons were gel purified
using the QIAquick Gel Extraction Kit (Qiagen, Germany) as per manufacturer protocol and sent to Inqaba Biotechnologies for direct
sequencing.
Analysis
Sequences were aligned and analysed using BioEdit freeware (http://www.mbio.ncsu.edu/BioEdit/bioedit.html). Sequences were exported into the
National Center for Biotechnology Information (NCBI) website’s Basic Local Alignment Search Tool (BLAST) database for species identification
of the isolates. A tree of relatedness was drawn using the neighbor-joining method (Saitou & Nei 1987) of molecular evolutionary genetics
analysis (MEGA4) freeware (http://www.megasoftware.net) (Tamura et al. 2007).
Isolation of Bartonella from human and animal blood specimens
A total of 382 HIV-positive patients attending the HIV clinic volunteered to take part in this study; 246 (64%) were women and 136 (36%) were men.
Ethylenediaminetetra-acetic acid blood samples (whole blood including plasma) were plated out after having been freeze-thawed from minus 20 °C.
Blood cultures from this sample group yielded no Bartonella spp. isolates. Approximately a third (130/382) of the blood cultures were
contaminated by fungal or environmental bacteria growth due to the prolonged incubation periods. In order to decrease the amount of contamination,
amphotericin B was included in the blood culture technique. To assess the effect of amphotericin B on the growth
of Bartonella, the B. henselae ATCC strain was inoculated into BHI broth with 5% (w/v) amphotericin B and plated out onto
fresh Bartonella media. Upon assessment, it was observed that although amphotericin B does reduce the amount of fungal contamination,
it also suppresses the growth of the control culture. The addition of amphotericin B was therefore discontinued. Due to the low circulating
bacterial load often observed in natural infection, the risk of suppression, although slight, was not one worth taking. If a culture became
contaminated with fungal growth, the culture was repeated. To ensure that Bartonella was not missed, any cultured organisms were
Gram-stained, and any Gram-negative pleomorphic bacilli were sub-cultured and Bartonella was excluded by PCR.A total of 42 clinically healthy volunteers were recruited to determine normal infection rates. Seven animal shelter staff and 35 National
Institute for Communicable Diseases’ staff members participated in this study. All specimens were freshly cultured onto
the Bartonella media. Only one of the 42 blood specimens illustrated growth morphologically consistent
with Bartonella spp. Unfortunately, the culture failed to grow on sub-culture. The seven animal shelter volunteers had daily exposure to many different animals. Bartonella spp. infections have been associated with
cats and dogs (Barnes et al. 2000; Ketring et al. 2004) and it was therefore expected that this group would be most likely to
demonstrate Bartonella bacteraemia. This was not the case in this study, but the sample size was inadequate. Dog (n = 179) blood specimens yielded no Bartonella culture isolates, but cat (n = 98) blood specimens yielded five
(approximately 5%) Bartonella culture isolates that were confirmed by PCR. It was concerning that there were so few cultured isolates and
that three of the five isolates obtained came from a single batch of seven feline blood samples received from the animal shelter, whereas
preceding batches had not yielded culture isolates. To exclude the possibility of cross-contamination within this batch, the blood samples were
re-cultured and subjected to PCR confirmation. A total of 124 rodent samples taken from Rattus norvegicus and Rattus rattus were cultured on Bartonella medium.
Bartonella was isolated from 16 (13%) rodents. All isolates were Gram-negative, pleomorphic bacilli with morphology consistent
with Bartonella. Primary isolation occurred between five and nine days, with sub-cultures growing within four to five days. At least two
different colony morphologies (Table 3) were observed for the rodent isolates, and these were later confirmed and identified by sequencing.
|
TABLE 3:
Two different colony morphologies observed for the rodent isolates.
|
Molecular detection and confirmation of Bartonella
A total of 21 culture isolates were confirmed as Bartonella by PCR. Amplicon sizes varied between 728 and 809 bp.
 |
FIGURE 1:
Molecular detection and confirmation of Bartonella.
|
|
Polymerase chain reaction of the HIV-positive population yielded a prevalence of 22.5% (86/382) (95% confidence; 18.5–27.1), whereas the
clinically healthy group had a prevalence of 9.5% (4/42) (95% confidence; 3.1–23.5). This is a significant difference (p-value: 0.05;
chi-square statistic: 3.818 with one degree of freedom) in the proportion of current infection for the two populations. This difference is unlikely
to have occurred through mere chance, although the limited healthy volunteer sample size is a major limitation. The feline blood tested by PCR indicated a 23.5% (23/98) (95% confidence; 15.8–33.3) Bartonella prevalence. This was significantly
different (p-value: 0.0002; chi-square statistic: 13.500 with one degree of freedom) to the culture prevalence (5%). Both test techniques
test for current infection; however, due to the fastidious nature of the bacteria, PCR is the far more efficient method for detection
of Bartonella spp. as it does not rely on the viability of the bacteria. The dog PCR prevalence was 9% (16/179) (95% confidence; 5.4–14.4), which is significantly lower than the prevalences in
felines (p-value: 0.0009; chi-square statistic: 11.053 with one degree of freedom) and rodents (p-value: 0.0001; chi-square
statistic: 14.419 with one degree of freedom). Rat bloods tested by PCR indicated 25% prevalence (31/124) (95% confidence; 17.9–33.7). There was a significant difference
(p-value: 0.0151; chi-square statistic: 5.907 with one degree of freedom) between PCR prevalence and culture prevalence (13%). When the
prevalence for rats was compared with that of the felines, there was no significant difference (p-value: 0.7918; chi-square statistic:
0.070 with one degree of freedom).
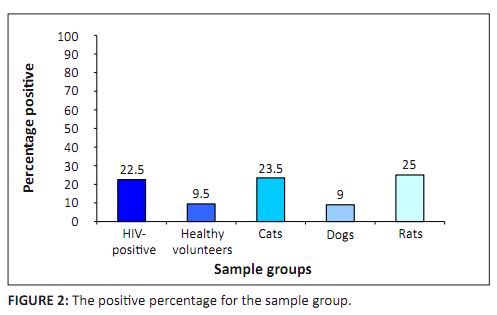 |
FIGURE 2:
The positive percentage for the sample group.
|
|
Deoxyribonucleic acid sequencing and analysis of Bartonella isolates
Comparison with GenBank (NCBI website) sequences showed that the rodent isolates ranged in percentage similarity from 97% – 99% to either the
recently named novel species candidatus ‘Bartonella thailandensis’ (RN24BJ; accession number: EF190333.1) or RN28BJ
(accession number: EF213776.1). Both isolates were described in a recent publication by Saisongkorh et al. (2009) where the rodents in
Beijing, China were tested for bartonellae. The rodent isolates were slightly more variable and a tree of relatedness was constructed (Figure 3). Contingent from the ITS data using parsimony
and distance methods illustrated two well-supported (more than 90% bootstrap values) clusters within the isolates. The first cluster placed RN24BJ
with 12 of the isolates (BART0272, BART0323, BART0268, BART0354, BART0379, BART0381, BART0312, BART0324, BART0359, BART0361, BART0357 and BART0358)
and the second group clustered RN28BJ with the remaining three isolates (BART0271, BART 0355 and
BART0377). Bartonella elizabethae (GenBank accession number: L35103)
and Bartonella grahamii (GenBank accession number: AJ269785) were used for comparison. Bartonella elizabethae was found to be
most similar to the rodent isolates from this study.
The five cat isolates were 99% – 100% similar to B. henselae URBHLIE 9 (accession number: AF312496.1). Primers (QHVE1 & QHVE3)
amplified a region consisting of 687 bp (excluding primers) for all the feline isolates. BART0480 and BART0483 were 100% identical to the B.
henselae URBHLIE-9 strain, and had only one nucleotide difference from B. henselae Houston-1 (accession number: L35101) strain at
position 98. BART0519 was 99% similar to URBHLIE-9 with a heterogenous nucleotide at position 285, where adenosine (A) or guanine (G) is equally
expressed. BART0519 and BART0484 were identical to each other and 99% similar to URBHLIE-9. One nucleotide difference was observed at position 660.
Bartonella remains one of the most difficult organisms to detect via blood or tissue culture. Frean et al. (2002) looked at the
prevalence of Bartonella in HIV-positive out-patients at several hospitals in Johannesburg and found a 10% PCR prevalence
of Bartonella. A conservative expectation for the present study was to find at least 10% culture-positive specimens
for Bartonella spp. Bartonella henselae was first isolated from the bloodstream of an AIDS patient (Regnery et al. 1992a). Severely immunocompromised
people are most at risk of contracting a Bartonella infection (Boulouis et al. 2005) and those with bacillary angiomatosis remain
bacteremic for a number of weeks (Koehler & Tappero 1993). HIV-infected patients with CD4+ cell counts of less than 50/mm3 are
more likely to develop bacillary angiomatosis lesions (Boulouis et al. 2005; Koehler & Tappero 1993), and as a precaution,
broad-spectrum antibiotic treatments are periodically prescribed for these patients. The effect of broad-spectrum antibiotics
on Bartonella infections lacks clarity. Research has been conducted on in vivo and in vitro antimicrobial
sensitivities of Bartonella to various drug classes. In vitro testing has shown bactericidal efficacy, but in vivo tests have
demonstrated mostly bacteristatic activity, with the exception of aminoglycosides which are bactericidal (Florin et al. 2008). The
bacteriostatic activity of the broad-spectrum drugs may be suppressing circulating bacteremia to undetectable levels for culture. It is probable
that the HIV-positive patients we tested were on a range of broad-spectrum antibiotics as prophylaxis against opportunistic infections.
Information regarding whether or not these patients were on the antibiotic treatment, the type of antibiotic treatment taken, the prescribed doses,
and duration of treatment administered was not collected at the time of specimen collection and is thus unavailable. It was speculated that this
would influence Bartonella isolation from the specimens. Published methodologies were followed for culture, including freeze-thawing of the specimens (La Scola & Raoult 1999). The methodologies were
later refined as it was discovered that inoculating fresh packed red-blood cell samples onto the Bartonella media yielded better results.
Although culture is regarded as the gold standard for detection, it is limited and problematic. The lack of cultured Bartonella in human
blood does not indicate that the prevalence is zero, since Bartonella spp. was detected by PCR. This study has shown an even higher Bartonella prevalence (22.5%) in HIV-positive out-patients than the 10% previously reported
(Frean et al. 2002). In immunocompromised individuals B. henselae infections are usually associated with exposure to cats and cat
fleas (Boulouis et al. 2005; Koehler & Tappero 1993). The highest prevalences found in this study were for cats (23.5%) and rats (25%).
These prevalences were not as high as some of the other reports published on the prevalence of bartonellae in animals. Dog and cat samples were
stored in the fridge at 4 °C for up to three weeks before being processed. It is hypothesised that Bartonella bacilli (if present in
the blood) became non-viable due to prolonged storage time. Supporting this hypothesis is the fact that isolates were obtained from blood
specimens that were processed within one week of collection. This is further supported by the fact that PCR detected Bartonella DNA
in the blood at a much higher rate than culture (23.5% vs 5%). An interesting finding for the present study was the isolation of B. henselae URBHLIE9 from all five culture-positive cat isolates. This
strain was previously isolated from the blood of a patient presenting with endocarditis and implies a strong link between humans and cats as
reservoirs for bartonellae (Houpikian & Raoult 2001). Polymerase chain reaction results indicate that there is a high prevalence of bartonellae in human and animal populations in Gauteng Province,
South Africa. More work is required to fully understand the extent of disease resulting from these Bartonella infections.
Limitations
Bartonella prevalence for HIV-positive patients in Gauteng was found to be much higher than previously reported. Based on the 22.5% (95%
confidence) PCR prevalence, it is speculated that approximately 1 in 4 HIV-positive patients is infected with Bartonella. Although most
patients are consistently subjected to broad-spectrum antibiotic treatment, the efficacy of treatment is not known since treatment efficacy varies
from patient-to-patient. The fact that 22.5% of the tested samples were PCR positive for Bartonella indicates that the bacteria are present
in the host. In vitro efficacy of antibiotic use against Bartonella has been limited (Florin et al. 2008). In most cases
antibiotics have had an inhibitory effect rather than a bactericidal one. The lack of cultured isolates from HIV-positive human samples could be
attributed to this inhibitory effect.Treatment is onerous as it has to be administered for prolonged periods to suppress infections, such as Bartonella infections. There is
limited information on the economic burden of Bartonella infections, since diseases caused by Bartonella are not notifiable.
Furthermore, due to the difficulty in diagnosis of Bartonella, infections often go undetected and untreated. Although there are various methodologies available for detection of Bartonella, each method has its own limitations. Most methods are
only used for research purposes and highly-skilled laboratory staff. The first test dealt with in this study is culture. Most microbiology
laboratories are proficient in this method and culture is generally regarded as the gold standard for most bacterial diagnoses. Culture
of Bartonella differs from most conventional bacteria culture as it is more labor-intensive. Fresh EDTA blood must be centrifuged and the
pellet is plated onto specialised media supplemented with rabbit blood. Isolation requires a prolonged incubation period from 7–21 days at
37 ºC under microaerophillic conditions. Most laboratories do not have the capacity to maintain these cultures for prolonged periods or
prevent contamination. Assuming the above conditions are met, there still remains the task of confirmation of the
culture as Bartonella. Bartonellae do not visibly metabolise compounds provided in rapid biochemical test panels. Bartonella spp. is
oxidase and catalase negative and stain faintly as Gram negative, slightly curved, bacilli. Small diagnostic laboratories often do not have the
means of confirming the culture isolate as Bartonella. Polymerase chain reaction has become a relatively affordable option when testing for bacterial diseases, although affordability is relative. A
skilled and competent operator performing a PCR may not have problems carrying out the test, particularly if it is a routine test, but if the
operator does not routinely perform the test (especially for Bartonella) the method is relatively labour-intensive and prone to
contamination. Much research has gone into developing nested PCR, high-tech real-time PCR and amplicon sequencing. These tests remain largely for
research purposes only. Staff would have to be trained; primers and other reagents would have to be readily available and expensive equipment
such as thermocyclers, electrophoresis power packs and tanks, UV light-boxes, software and computers would have to be purchased and maintained.
Recommendations
It is recommended that operator-friendly tests such as the IFA become more affordable for use in laboratories. Perhaps an ELISA as previously
described (Bergmans et al. 1997; Vermeulen et al. 2007) would be a more cost effective method. Control culture is relatively easy to
grow and the antigens of the culture can be sonicated from the growth and consequently be used to coat the microtitre wells. This method can be
optimised and made ‘in-house’ thus mitigating the need for expensive commercially available kits. Bartonella testing is
especially required for immunocompromised patients presenting with clinical symptoms and histories.Training and out-reach programs directed towards doctors and the generally public should be carried out to ensure the indicative symptoms are
recognised, and reported when presenting for medical assistance. Pest control and fleas control methods should be advised and implementable
solutions should also be discussed.
In this study, results were within global trends for Bartonella prevalence in both human and animal populations. The objectives of the study
were met and a better understanding of Bartonella prevelance in human and animal populations in Gauteng, South Africa is now available. This
study has confirmed that the primary concern is human health, particularly for immunocompromised people who are at higher risk of contracting
various opportunistic infections including Bartonella. Bartonella is prevalent in human and animal populations in Gauteng, South Africa, and could be responsible for a number of unresolved or
misdiagnosed diseases in immunocompromised patients. Although the broad-spectrum antibiotics offered to HIV-positive patients in particular maybe
suppressing the infection, the treatment would have to be constant and there are possible side-effects that could cause the patient more
discomfort. It is important to emphasise that immunocompromised patients be very careful when handling domestic animals. Furthermore, attempts should be made to decrease the risk of rat exposure. By keeping food in sealed containers off the floor, and ensuring
proper sanitation in and around the home is followed, the household rodent population would be largely reduced. Too little is known about Bartonella and further research is required to fully understand the extent of disease related to this emergent
pathogen.
It is with gratitude that we would like to acknowledge the following; firstly the staff of SBPRL – CEZD for their support and assistance with
the project, Mr Justice Matebula (Laboratory aid) who assisted with the translations and explanations of the informed consent in the recruitment of
volunteer participants, the HIV-clinic councilors and phlebotomists for their assistance with translations, explanations and recruitment of
HIV-positive volunteers as well as specimen collection, and to the staff of NICD and the animal shelter (wished to remain anonymous)
for ‘bleeding’ for the cause. The animal shelter is furthermore thanked for assisting us with obtaining the cat and dog specimens. A
special thank you is owed to Mr Lee Ashford and his enthusiastic staff at ‘Scientific Supakill’ pest-control company for providing us
with the live rodents. And finally thank you to the NICD/NHLS for their support and use of facilities in this study.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this paper.
Birtles, R.J., Laycock, G., Kenny, M.J., Shaw, S.E. & Day, M.J., 2002, ‘Prevalence of Bartonella species causing bacteraemia in
domesticated and companion animals in the United Kingdom’, Veterinary Record 151, 225–229.
http://dx.doi.org/10.1136/vr.151.8.225,
PMid:12219899
Bonilla, D.L., Kabeya, H., Henn, J., Kramer, V.L. & Kosoy, M.Y., 2009, ‘Bartonella quintana in body lice and head lice from
homeless persons, San Francisco, California, USA’, Emerging Infectious Diseases 15, 912–915.
http://dx.doi.org/10.3201/eid1506.090054,
PMid:19523290
Boulouis, H.J., Chang, C.C., Henn, J.B., Kastenc, R.W. & Chomel, B.B., 2005, ‘Factors associated with the rapid emergence of
zoonotic Bartonella infections’, Veterinary Research 36, 383–410.
http://dx.doi.org/10.1051/vetres:2005009,
PMid:15845231
Branley, J., Wolfson, C., Waters, P., Gottlieb, T. & Bradbury, R., 1996, ‘Prevalence of Bartonella henselae bacteremia, the
causative agent of cat scratch disease, in an Australian cat population’, Pathology 28, 262–265.
http://dx.doi.org/10.1080/00313029600169124,
PMid:8912359
Breitschwerdt, E.B.,Maggi, R.G.,Duncan, A.W.,Nicholson, W.L.,Hegarty, B.C. & Woods, C.W., 2007, ‘Bartonella species in blood of
immunocompetent persons with animal and arthropod contact’, Emerging Infectious Diseases 13, 938–941.
http://dx.doi.org/10.3201/eid1306.061337,
PMid:17553243
Chomel, B.B., Boulouis, H.J., Maruyama, S. & Breitschwerdt, E.B., 2006, ‘Bartonella spp. in pets and effect on human
health’, Emerging Infectious Diseases 12, 389–394.
http://dx.doi.org/10.3201/eid1203.050931,
PMid:16704774
Duncan, A.W., Maggi, R.G. & Breitschwerdt, E.B., 2007, ‘A combined approach for the enhanced detection and isolation
of Bartonella species in dog blood samples: Pre-enrichment liquid culture followed by PCR and subculture onto agar plates’,
Journal of Microbiological Methods 69, 273–281.
http://dx.doi.org/10.1016/j.mimet.2007.01.010,
PMid:17346836
Florin, T.A., Zaoutis, T.E. & Zaoutis, L.B., 2008, ‘Beyond cat scratch disease: Widening spectrum of Bartonella henselae
infection’, Pediatrics 121, 1413–1425.
http://dx.doi.org/10.1542/peds.2007-1897,
PMid:18443019
Frean, J., Arndt, S. & Spencer, D., 2002, ‘High rate of Bartonella henselae infection in HIV-positive outpatients in
Johannesburg, South Africa’, Transactions of the Royal Society of Tropical Medicine and Hygiene 96, 549–550.
http://dx.doi.org/10.1016/S0035-9203(02)90437-2 Greub, G. & Raoult, D., 2002, ‘Bartonella: New explanations for old diseases’, Journal of Medical
Microbiology 51, 915–923.
PMid:12448674
Heller, R., Artois, M., Xemar, V., De Briel, D., Gehin, H., Jaulhac, B. et al., 1997, ‘Prevalence of Bartonella
henselae and Bartonella clarridgeiae in stray cats’, Journal of Clinical Microbiology 35, 1327–1331.
PMid:9163438
Houpikian, P. & Raoult, D., 2001, ‘16S/23S rRNA intergenic spacer regions for phylogenetic analysis, identification, and
subtyping of Bartonella species’, Journal of Clinical Microbiology 39, 2768–2778. Jacomo, V., Kelly, P.J. & Raoult, D., 2002, ‘Natural history of Bartonella infections (an exception to Koch’s
postulate)’, Clinical and Diagnostic Laboratory Immunology 9, 8–18.
PMid:11777823
Kamrani, A., Parreira, V.R., Greenwood, J. & Prescott, J.F., 2008, ‘The prevalence of Bartonella, hemoplasma,
and Rickettsia felis infections in domestic cats and in cat fleas in Ontario’, The Canadian Journal of Veterinary
Research 72, 411–419.
PMid:19086373
Kelly, P.J., Matthewman, L.A., Hayter, D., Downey, S., Wray, K., Bryson, N.R. & Raoult, D.,
1996, ‘Bartonella (Rochalimaea) henselae in southern Africa – evidence for infections in domestic cats and
implications for veterinarians’, Journal of the South African Veterinary Association 67, 182–187.
PMid:9284029
Kim, C.M., Kim, J.Y., Yi, Y.H., Lee, M.J., Cho, M.R., Shah, D.H. et al., 2005, ‘Detection of Bartonella species from ticks,
mites and small mammals in Korea’, Journal of Veterinary Science 6(4), 327–334.
PMid:16293997
Kim, Y.S., Seo, K.W., Lee, J.H., Choi, E.W., Lee, H.W., Hwang, C.Y. et al., 2009, ‘Prevalence of Bartonella
henselae and Bartonella clarridgeiae in cats and dogs in Korea’, Journal of Veterinary Science 10(1),
85–87.
http://dx.doi.org/10.4142/jvs.2009.10.1.85,
PMid:19255530
Koehler, J.E. & Tappero, J.W., 1993, ‘Bacillary angiomatosis and bacillary peliosis in patients infected with human immunodeficiency
virus’, Clinical Infectious Diseases 17, 612–624.
http://dx.doi.org/10.1093/clinids/17.4.612,
PMid:8268340
Koehler, J.E., Sanchez, M.A., Tye, S., Garrido-Rowland, C.S., Chen, F.M., Maurer, T. et al., 2003, ‘Prevalence
of Bartonella infection amoung human immunodeficiency virus infected patients with fever’, Clinical Infectious Diseases 37,
559–566.
http://dx.doi.org/10.1086/375586,
PMid:12905141
La Scola, B. & Raoult, D., 1999, ‘Culture of Bartonella quintana and Bartonella henselae from Human Samples: A
5-Year Experience (1993 to 1998)’, Journal of Clinical Microbiology 37, 1899–1905. Maggi, R.G., Kosoy, M., Mintzer, M. & Breitschwerdt, E.B., 2009, ‘Isolation of Candidatus Bartonella melophagi from Human
Blood’, Emerging Infectious Diseases 15(1), 66–68.
http://dx.doi.org/10.3201/eid1501.081080,
PMid:19116054
Márquez, F.J., Rodríguez-Liébana, J.J., Pachón-Ibáñez, M.E., Docobo-Pérez, F.,
Hidalgo-Fontiveros, A., Bernabeu-Wittel, M. et al., 2008, ‘Molecular Screening of Bartonella Species in Rodents from South
Western Spain’, Vector-Borne and Zoonotic Diseases 8(5), 695–700.
http://dx.doi.org/10.1089/vbz.2007.0257,
PMid:18620508
Maurin, M., Roux, V., Stein, A., Ferrier, F., Viraben, R. & Raoult, D., 1994, ‘Isolation and characterisation by immunofluorescence,
sodium dodecyl sulfate-polyacrylamide gel electrophoresis, western blot, restriction fragment length polymorphism-PCR, 16srRNA gene sequencing,
and pulsed-field gel electrophoresis of Rochalimaea quintana from a patient with bacillary angiomatosis’, Journal of Clinical
Microbiology 32, 1166–1171.
PMid:7519628
Maurin, M. & Raoult, D., 1996, ‘Bartonella (Rochalimaea) quintana infections’, Clinical Microbiology
Reviews 9, 273–292.
PMid:8809460
Pérez-Martínez, L., Venzal, J.M., González-Acuña, D., Portillo, A., Blanco, J.R. & Oteo, J.A.,
2009, ‘Bartonella rochalimae and other Bartonella spp. in Fleas, Chile’, Emerging Infectious Diseases 15(7),
1150–1152.
http://dx.doi.org/10.3201/eid1507.081570,
PMid:19624952
Pons, I., Sanfeliu, I., Nogueras, M.M., Sala, M., Cervantes, M., Amengual, M.J. et al., 2008, ‘Seroprevalence
for Bartonella spp. infection in HIV patients in Catalonia, Spain’, BioMed Central Infectious Diseases 8, 58.
http://dx.doi.org/10.1186/1471-2334-8-58,
PMid:18452613
Pretorius, A., Beati, L. & Birtles, R.J., 2004, ‘Diversity of bartonellae associated with small mammals inhabiting Free State province,
South Africa’, International Journal of Systematic and Evolutionary Microbiology 54, 1959–1967.
http://dx.doi.org/10.1099/ijs.0.03033-0,
PMid:15545418
Rampersad, J.N., Watkins, J.D., Samlal, M.S., Deonanan, R., Ramsubeik, S. & Ammons, D.R., 2005, ‘A nested-PCR with an Internal
Amplification Control for the detection and differentiation of Bartonella henselae and B. clarridgeiae: An examination of cats in
Trinidad’, BioMed Central Infectious Diseases 5, 63–68.
http://dx.doi.org/10.1186/1471-2334-5-63,
PMid:16098227
Regnery, R.L., Anderson, B.E., Clarridge, J.E., Rodriguez-Barradas, M.C., Jones, D.C. & Carr, J.H., 1992, ‘Characterization of a
novel Rochalimaea species, R. henselae sp. Nov., isolated from blood of a febrile, human immunodeficiency virus-positive
patient’, Journal of Clinical Microbiology 30, 265–274.
PMid:1371515
Rolain, J.M., Locatelli, C., Chabanne, L., Davoust, B. & Raoult, D., 2004, ‘Prevalence of Bartonella
clarridgeiae and Bartonella henselae in domestic cats from France and detection of the organisms in erythrocytes by
Immunofluorescence’, Clinical and Diagnostic Laboratory Immunology 11(2), 423–425.
PMid:15013998
Roux, V. & Raoult, D., 1995, ‘Inter- and intraspecies identification of Bartonella (Rochalimaea) species’,
Journal of Clinical Microbiology 33, 1573–1579.
PMid:7650189
Sander, A., Buhler, C., Pelz, K., Von Cramm, E. & Bredt, W., 1997, ‘Detection and identification of two
Bartonella henselae variants in domestic cats in Germany’, Journal of Clinical Microbiology 35(3), 584–587.
PMid:9041393
Seki, N., Sasaki, T., Sawabe, K., Sasaki, T., Matsuoka, M., Arakawa, Y. et al., 2006, ‘Epidemiological Studies on Bartonella
quintana Infections among Homeless People in Tokyo, Japan’, Japanese Journal of Infectious Diseases 59(1), 31–35.
PMid:16495631
Winoto, I.L., Goethert, H., Ibrahim, I.N., Yuniherlina, I., Stoops, C., Susanti, I. et al., 2005, ‘Bartonella species in
rodents and shrews in the Greater Jakarta Area’, Southeast Asia Journal of Tropical Medicine and Public
Health 36(6), 1523–1529.
PMid:16610656
Wong, M.T., Thornton, D.C., Kennedy, R.C. & Dolan, M.J., 1995, ‘A chemically defined liquid medium that supports primary isolation
of Rochalimae (Bartonella) henselae from blood and tissue specimens’, Journal of Clinical
Microbiology 33, 742–744.
PMid:7538511
Yabsley, M.J., McKibben, J., Macpherson, C.N., Cattan, P.F., Cherry, N.A., Hegarty, B.C. et al., 2008, ‘Prevalence
of Ehrlichia canis, Anaplasma platys, Babesia canis vogeli, Hepatozoon canis, Bartonella vinsonii
berkhoffii, and Rickettsia spp. in dogs from Grenada’, Veterinary Parasitology 151, 279–285.
http://dx.doi.org/10.1016/j.vetpar.2007.11.008,
PMid:18160223
Ying, B., Kosoy, M.Y., Maupin, G.O., Tsuchiya, K.R. & Gage, K.L., 2002, ‘Genetic and ecologic characteristics of Bartonella
communities in rodents in Southern China’, American Journal of Tropical Medicine and Hygiene 66(5), 622–627.
PMid:12201602
|
